
RNAi Explorer |
|
siRNA Design Strategies: The strategy for siRNA design is based on our present understanding of the biochemical mechanisms involved in RNA interference and, in particular, structural features that allow the antisense-strand of the siRNA duplex to be more efficiently incorporated into the RNA-induced silencing complex (RISC). Default parameters: (N19)TT 1. Low to medium GC content (30-50%).2. Absence of internal repeats or palindromes. 3. Presence of an A at position 3 of the sense strand. 4. Presence of A at position 19 of the sense strand. 5. Absence of G or C at position 19 of the sense strand. 6. Presence of U at position 10 of the sense strand. 7. Absence of a G at position 13 of the sense strand. 8. At least 3 A/Us at positions 15-19 of the sense strand. 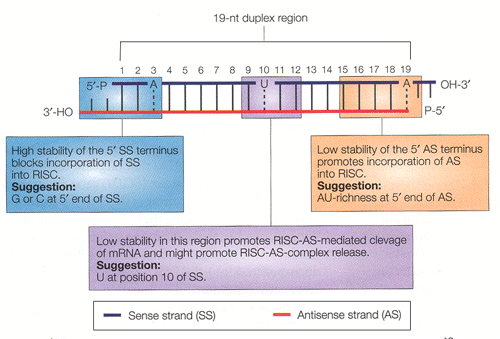
AA(19mer) The default criteria selects target sequences of 21 nucleotides that begin with AA and are located within a region of the coding sequence that is within 50-100 nucleotides of the AUG start codon and within 50-100 nucleotides from the termination codon. The presence of AA at the start of the sequence allows for the use of dTdT at the 3'-end of the antisense sequence. The sense strand can also be synthesized with dTdT at the 3' end, because only the antisense strand is involved in target recognition. The use of dTdT reduces the cost of synthesis and also makes the siRNA duplex more resistant to exonuclease activity. Because a number of reports have demonstrated that the presence of AA at the beginning of the target sequence is not an absolute requirement, the selection program includes the option to search for sequences that begin with other nucleotide pairs. Figure 1: Nature Genetics May 2004, Volume 5 No 5. Author : Vivek MittalGC Content The G-C content of the sequence is also used as a condition for selecting target sequences. Ideally the GC content will be less than 50%, although successful gene silencing has been reported with siRNAs that have G-C contents between 50 and 60%. The default parameter selects for a G-C content in the 40-50% range, however, options are available that allow for selection over wider ranges. |
|
Stretches of Nucleotide Repeats The default mode avoids sequences with repeats of three or more G's or C's, as their presence initiates intra-molecular secondary structures preventing effective siRNA silencing hybridization. As an option, repeat stretches of A's and T's can also be eliminated, as they tend to reduce the specificity of the target sequence. If possible, this option is highly recommended. |
|
Blast Search Once a target sequence has been chosen, a BLAST search is initiated to ensure that your target sequence is not homologous to other gene sequences. Target sequences that have more than 15 contiguous bases pairs of homology to other genes in the NCBI database are eliminated. |
|
What is siRNA? Recent advances in molecular biology have shown that gene expression can be effectively silenced in a highly specific manner through the addition of double stranded RNA (dsRNA) (1-3). The term RNA interference (RNAi) was created to describe this phenomenon and, while the mechanism was originally observed in plants and later in the worm Caenorhabditis elegans, subsequent studies have shown that RNAi is present in a wide variety of eukaryotic organisms including mammals (4-6). For the most part, it is believed that RNAi serves as an antiviral defense mechanism although there is preliminary evidence that it also plays a role in the formation and maintenance of heterochromatin during mitosis and meiosis (7,8). Once dsRNA enters the cell, it is cleaved by an RNase III –like enzyme, Dicer, into double stranded small interfering RNAs (siRNA) 21-23 nucleotides in length that contain 2 nucleotide overhangs on the 3' ends (9-11). In an ATP dependent step, the siRNAs become integrated into a multisubunit protein complex, commonly known as the RNAi induced silencing complex (RISC), which guides the siRNAs to the target RNA sequence (12). At some point the siRNA duplex unwinds, and it appears that the antisense strand remains bound to RISC and directs degradation of the complementary mRNA sequence by a combination of endo and exonucleases (13). Preliminary studies in mammalian systems using long dsRNAs to initate the RNAi response failed because they led to the induction of a non-specific Type I interferon response that produced extensive changes in protein expression and eventually resulted in cell death (14,15). Subsequent studies, however, using synthetic, short double-stranded RNAs that mimic the siRNAs produced by the enzyme dicer, sequence specific gene silencing could be achieved in mammalian cells without inducing the interferon response (6,16). siRNA technology is now extensively recognized as a powerful tool for the specific suppression of gene expression and is presently being used by researchers in a wide range of disciplines for the assessment of gene function.
|
|
References 1. Fire,A., Xu,S., Montgomery, M.K., Kostas, S.A., Driver, S.E. and Mello, C.C. (1998) Nature, 391, 806 811. 2. Napoli, C., Lemieux, C. & Jorgensen, R. (1990) Plant Cell 2, 279 289. 3. Hannon, G.J. (2002) RNA interference. Nature, 418, 244 251 4. Billy,E., Brondani,V., Zhang,H., Muller,U. and Filipowicz,W. (2001) Proc. Natl. Acad. Sci. USA, 98, 14428 14433 5. Paddison, P.J., Caudy, A.A. and Hannon, G.J. (2002) Proc. Natl Acad. Sci. USA, 99, 1443 1448 6. Elbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K. and Tuschl,T. (2001) Nature, 411, 494 498 7. Volpe TA, Kidner C, Hall IM, Teng G, Grewal SI, Martienssen RA. (2002) Science.,297, 1833-1837. 8. Allshire R. (2002) Science, 297, 1818-1819. 9. Elbashir, S.M., Lendeckel, W. and Tuschl, T. (2001) Genes Dev., 15, 188 200. 10. Bernstein, E., Caudy, A.A., Hammond, S.M. and Hannon, G.J. (2001) Nature, 409, 363 366. 11. Hammond, S.M., Bernstein, E., Beach, L. and Hannon, G.J. (2000) Nature, 404, 293 296. 12. Nykanen, A., Haley, B. and Zamore, P.D. (2001) Cell, 107, 309 321. 13. Martinez, J., Patkaniowska, A., Urlaub, H., Luhrmann, R. and Tuschl, T. (2002) Cell, 110, 563 574. 14. Caplen, N.J., S. Parrish, F. Imani, A. Fire and R.A. Morgan. 2001. Proc Natl Acad Sci U S A 98:9742-9747. 15. Ullu E, Djikeng A, Shi H, Tschudi C., 2002, Philos Trans R Soc Lond B Biol Sci. 357, 65-70. 16. Ui-Tei, K., S. Zenno, Y. Miyata and K. Saigo, 2000, FEBS Lett 479:79-82. 18. Mittal V (2004) Nature Reviews 5:355-365. |


 RNAi - Technical Information
RNAi - Technical Information